Article Text

Abstract
Ferulic acid, a bacterial metabolite of anthocyanins, seems likely to be a primary mediator of the health benefits associated with anthocyanin-rich diets, and has long been employed in Chinese cardiovascular medicine. In rodent studies, it has exerted wide-ranging antioxidant and anti-inflammatory effects, the molecular basis of which remains rather obscure. However, recent studies indicate that physiologically relevant concentrations of ferulic acid can boost expression of Sirt1 at mRNA and protein levels in a range of tissues. Sirt1, a class III deacetylase, functions to detect a paucity of oxidisable substrate, and in response works in various ways to promote cellular survival and healthful longevity. Sirt1 promotes ‘cell cleansing’ and cell survival by boosting autophagy, mitophagy, mitochondrial biogenesis, phase 2 induction of antioxidant enzymes via Nrf2, and DNA repair—while inhibiting NF-kB-driven inflammation, apoptosis, and cellular senescence, and boosting endothelial expression of the protective transcription factor kruppel-like factor 2. A deficit of the latter appears to mediate the endothelial toxicity of the SARS-CoV-2 spike protein. Ferulic acid also enhances the activation of AMP-activated kinase (AMPK) by increasing expression and activity of its activating kinase LKB1—whereas AMPK in turn amplifies Sirt1 activity by promoting induction of nicotinamide phosphoribosyltranferase, rate-limiting for generation of Sirt1’s obligate substrate NAD+. Curiously, AMPK acts by independent mechanisms to potentiate many of the effects mediated by Sirt1. Hence, it is proposed that ferulic acid may exert complementary or synergistic health-promoting effects when used in conjunction with clinically useful AMPK activators, such as the nutraceutical berberine. Additional nutraceuticals which might have potential for amplifying certain protective effects of ferulic acid/berberine are also discussed.
- carotid artery diseases
- coronary angiography
- echocardiography
Data availability statement
No data are available.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Upregulation of Sirt1 by ferulic acid
Ferulic acid is a bacterial metabolite of anthocyanins that may be a key mediator of the health-protective effects of anthocyanin-rich diets documented in epidemiology and rodent studies.1 It may also mediate the physiological effects of nutraceuticals such as pycnogenol and elderberry that provide anthocyanins.2 3 Various conjugated forms of ferulic acid, of differing degrees of bioavailability, are also found in many natural plant-based foods. In rodents, ferulic acid displays potent anti-inflammatory and antioxidant effects, associated with increased activity of the Nrf2 transcription factor—driver of the phase 2 antioxidant response—and decreased activity of the proinflammatory NF-kB transcription factor.1 The mechanistic basis of these effects requires further clarification. Clinically, oral or parenteral sodium ferulate has long been employed in Chinese medicine for a range of cardiovascular applications, and a recent Thai clinical study found that, at 1000 mg daily, ferulic acid improves serum lipid profile, reduces markers of oxidative stress, and decreases C reactive protein by about one-third.4 5
An important clue to the basis of ferulic acid’s versatile protective effects may have been provided by recent studies reporting that, in low micromolar concentrations that are readily achievable with oral supplementation and likely by ingestion of anthocyanin-rich diets, ferulic acid upregulates expression of Sirt1 at both the mRNA and protein level. This effect has been demonstrated in chondrocytes, skeletal muscle fibres, testes, neural stem cells and hepatocytes, and hence may be broad in scope.6–11 Presumably, ferulic acid either upregulates transcription of the Sirt1 gene or prolongs the half-life of Sirt1 mRNA. Sirt1 is a class III deacetylase that functions to detect a paucity of oxidisable substrate—as during caloric restriction; it responds to this signal by deacetylating a number of target proteins, thereby mediating homeostatic responses to perceived energy deficit.12 Sirt1 uses NAD+ as an obligate substrate; when the NAD+/NADH ratio is high—as it is when oxidisable fuel is low—Sirt1 activity increases.
The discovery that ferulic acid is a Sirt1 inducer may offer important insight into its effects on Nrf2 and NF-kappaB activities. Sirt1 promotes antioxidant phase 2 induction by deacetylating and enhancing the transcriptional activity of Nrf213–15; this effect likely contributes to the phase 2-inducing activity of ferulic acid, although activation of Erk1/2 is also thought to contribute in this regard.16–18 Ferulic acid-mediated induction of Sirt1 would also be expected to combat inflammation by reversing the acetylation of p65 on Lys310 required for optimal transactivation activity of the NF-kappaB complex.15 19 There is also evidence that Sirt1 may act more proximally in proinflammatory signalling by suppressing activating phosphorylation and ubiquitination of transforming growth factor-beta activated kinase 1 (TAK1).20 This presumably reflects the fact that activated Sirt1 binds directly to TAK1. It is not yet clear, however, whether this effect of Sirt1 reflects reversal of an activating acetylation of TAK1. Indeed, certain acetylations of TAK1 may reduce its proinflammatory activity.21 In any case, Sirt1-mediated inhibition of TAK1 could be expected to prevent TAK1-mediated activation of NF-kappaB and the stress activated MAP kinases p38 and JNK, which collaborate in inducing expression of many proinflammatory cytokines and enzymes. Other intriguing research shows that upregulated expression of the MyD88 coupling factor negates the inhibitory impact of ferulic acid on proinflammatory signalling, for reasons that remain unclear.1
Sirt1 and AMP-activated kinase collaborate in promoting autophagy, mitophagy and mitochondrial biogenesis
Of particular interest is the fact that Sirt1 and AMP-activated kinase (AMPK) exert mutually reinforcing effects. Sirt1 deacetylates and thereby enhances the stability and activity of LKB1, an enzyme which activates AMPK via phosphorylation.22 Conversely, AMPK promotes Sirt1 activity by upregulating expression of nicotinamide phosphoribosyltransferase (NAMPT), which is rate-limiting for re-generation of NAD+, a substrate for Sirt1; NAMPT in the process also lowers cellular levels of nicotinamide, an inhibitor of Sirt1 activity.23–26 These considerations suggest that there might a complementary or synergistic interaction between ferulic acid and drug or nutraceuticals that activate AMPK. In this regard, the antidiabetic activities of the drug metformin and the nutraceutical berberine is thought to stem in whole or in part by activation of AMPK. These agents, by modestly boosting cellular AMP levels, enable the activating phosphorylation of AMPK via LKB1.27 28 Hence, ferulic acid and metformin or berberine would be expected to exert complementary effects on AMPK activation. In this regard, a synergism between ferulic acid and metformin has been reported in diabetic rats.29 30 These considerations encourage the proposal that nutraceutical regimens providing both ferulic acid and berberine might have intriguing potential for health promotion. In particular, it is intriguing to note that Sirt1 and AMPK (which likewise functions as a detector of energy deficit, AMP and ADP enhance its activity) may collaborate in the promotion of autophagy, mitophagy and mitochondrial biogenesis (MB)—processes which jointly promote ‘clean’ cells by disposing of damaged proteins and organelles, while insuring that mitochondria function efficiently and generate only modest levels of oxidants.
Sirt1 and AMPK collaborate in complementary ways in promoting autophagy, which in moderation is thought to promote healthful ageing at the cellular level.31 Sirt1 acts in this regard by deacetylating various factors which in their deacetylated forms participate in the formation of autophagosomes, as well as via a deacetylation of FOXO1 transcription factor that enables transcription of the gene coding for Rab7, a G protein essential for autophagosome/lysosome fusion.32–34 In addition, Sirt1 promotes mitophagy by increasing expressions of Parkin and Pink1, which tag dysfunctional mitochondria for autophagic destruction.35–37 AMPK promotes autophagy by conferring an activating phosphorylation on ULK1 while suppressing the antiautophagic activity of mTORC1.38–40
While Sirt1 and AMPK promote autophagic disposal of ageing mitochondria (mitophagy), they concurrently collaborate to promote MB via activating post-translational modifications of PPAR-γ coactivator-1α (PGC-1α); the latter promotes transcription of a number of genes coding for proteins required for MB.41–43 Phosphorylation of PGC-1α by AMPK appears to be a prerequisite for subsequent deacetylation by Sirt1.44 Sirt1 also promotes MB via its upregulatory impact on Nrf2 activity; the latter increases expression of nuclear regulatory factor-1 which, via interaction with PGC-1α, transcriptionally upregulates expression of several key proteins required for transcription and replication of mitochondrial DNA.45 46 And, finally, Sirt1’s downregulatory impact on NF-kappaB activity likewise aids MB, as NF-kappaB opposes the synthesis and coactivation activity of PGC-1α.47 48 MB, coupled with mitophagy, maintains mitochondria in a functionally youthful state, such that they are efficient at ATP generation and less prone to induce oxidative stress. Sirt1, as a detector of inefficient cellular bioenergetics, functions homeostatically to boost production of mitochondria so as to help insure an adequate ATP supply.
By promoting a complex between deacetylated FOXO3a and PCG-1α, as well as by Nfr2 activation, Sirt1 increases expression of manganese-dependent superoxide dismutase and other enzymes that provide antioxidant protection to mitochondria.49 And Sirt1/AMPK-mediated activation of PGC-1α also decreases mitochondrial oxidant production by inhibiting expression of dynamin-related protein 1, a promoter of mitochondrial fission.50 Another consequence of PGC-1α activation is increased expression of Sirt3; this acts to minimise mitochondrial oxidant stress, both by modulating nuclear FOXO3a activity, and by acting as an intra-mitochondrial deacetylase.51–53 Hence, Sirt1/AMPK activities help to insure that newly synthesised mitochondria remain structurally sound and do not contribute unduly to cellular oxidant load.
Sirt1 also promotes cellular integrity by aiding the efficiency of DNA repair.54 This effect tends to prevent cell senescence by aiding maintenance of telomere length.55 Sirt1 likewise tends to oppose apoptosis—both via promotion of DNA repair, and by deacetylating and hence inactivating p53.54–56 Sirt1 modulation of FOXO3a opposes apoptosis by inhibiting FOXO3a-mediated transcription of the genes coding for the proapoptotic factors Bim and PUMA.57–59 Hence, Sirt1 activity aids cell survival under stress by aiding DNA repair and promoting autophagy, while discouraging apoptotic cell death. These effects would tend to suppress tumour initiation, but could aid tumour promotion by aiding the survival of DNA-damaged cells; stimulation of Sirt1 would presumably be contraindicated in the context of cancer chemotherapy.60 Downregulation of apoptosis would be particularly protective for tissues that do not readily regenerate, such as the heart and central nervous system. The anti-apoptotic effects of Sirt1, coupled with its anti-inflammatory and antioxidant activities, make Sirt1 a promising target for prevention of neurodegenerative disorders and of congestive heart failure.61 62
Sirt1/AMPK promote induction of kruppel-like factor 2, crucial for healthy endothelial function
The endothelial transcription factor kruppel-like factor 2 (KLF2) is induced by pulsatile shear stress (PSS), and exerts various anti-inflammatory and anti-thrombotic effects, while boosting expression of endothelial nitric oxide (NO) synthase.63 64 PSS also enhances the activity of AMPK while increasing the stability of Sirt165–67; moreover, both AMPK and Sirt1 have been reported to increase KLF2 at the transcriptional level via stimulation of the mitogen-activated protein kinase 5/myocyte enhancing factor 2 (MEF2) signalling pathway.66 68–71 AMPK also phosphorylates and induces nuclear export of HDAC5, which can interact with MEF2 to block its transcriptional activity.72 73 And, via deacetylation of NF-kB, Sirt1 may oppose a suppressive effect of NF-κB on transcription of KLF2 and on KLF2-mediated transcription.74 These findings suggest that a ferulic acid/berberine regimen may collaborate in promoting healthful endothelial function via KLF2 induction. The impact of Sirt1 activity on KLF2 may be pertinent to the use of sodium ferulate in Chinese cardiovascular medicine; analogously, AMPK-mediated KLF2 induction may help to rationalise the favourable influence of metformin treatment on cardiovascular risk in diabetics.4 75 76
In this regard, it is topical to note that exposure of endothelial cells to serum from patients with COVID-19 or to SARS-CoV-2 spike protein diminishes KLF2 expression, possibly via enhanced proteasomal degradation.77 78 Moreover, enhancing KLF2 expression via transfection, bortezomib, or statins was found to suppress the proinflammatory, prothrombotic effects of COVID-19 serum or of spike protein on endothelial cells.77 78 Hence, ferulic acid/berberine may have potential for controlling the endotheliopathy associated with COVID-19 or SARS-CoV-2 spike protein exposure (as via mRNA vaccines) by upregulation of KLF2. The upregulatory effect of statins on KLF2 expression has been traced to inhibition of protein prenylation and consequent inhibition of Rho activation.74 79 80 It is reasonable to suspect that modulation of KLF2 is pertinent to the reported favourable impacts of concurrent metformin or statin therapy on COVID-19 prognosis.81 82
AMPK can also boost Nrf2 activity
In at least certain contexts, AMPK can upregulate Nrf2-mediated induction of phase 2 proteins, in a manner that is independent of Sirt1.83–86 In cells which partially rely on insulin receptor substrate-1 (IRS-1) for mitogenic signalling, AMPK can boost Akt activation through inhibition of the mTORC1/p70S6 kinase pathway; the latter kinase confers phosphorylations on IRS-1 that reduce its stability.87 Akt, in turn, via deactivating phosphorylation of glycogen synthase kinase-3b (GSK3b), enhances the stability and nuclear localisation of Nrf2, aiding its transcriptional activity.88 89 GSK3b is capable of conferring an activating phosphorylation on Fyn kinase; this causes Fyn kinase to migrate to the nucleus and phosphorylate Nrf2, which induces its export from the nucleus and accelerates its proteasomal degradation. Hence, AMPK mediated Akt activation abrogates the inhibitory impact of GSK3b/Fyn kinase signalling on Nrf2 activity. Other recent studies show that AMPK can directly phosphorylate Nrf2; while these phosphorylations per se do not alter Nrf2 location or stability, they increase the ability of Nrf2 to promote transcription of some (including heme oxygenase-1) but not all Nrf2 target genes.90 In addition, AMPK-mediated phosphorylation of the transcriptional regulator Bach1 destabilises it, reducing its ability to inhibit transcription of certain Nrf2-responsive genes.
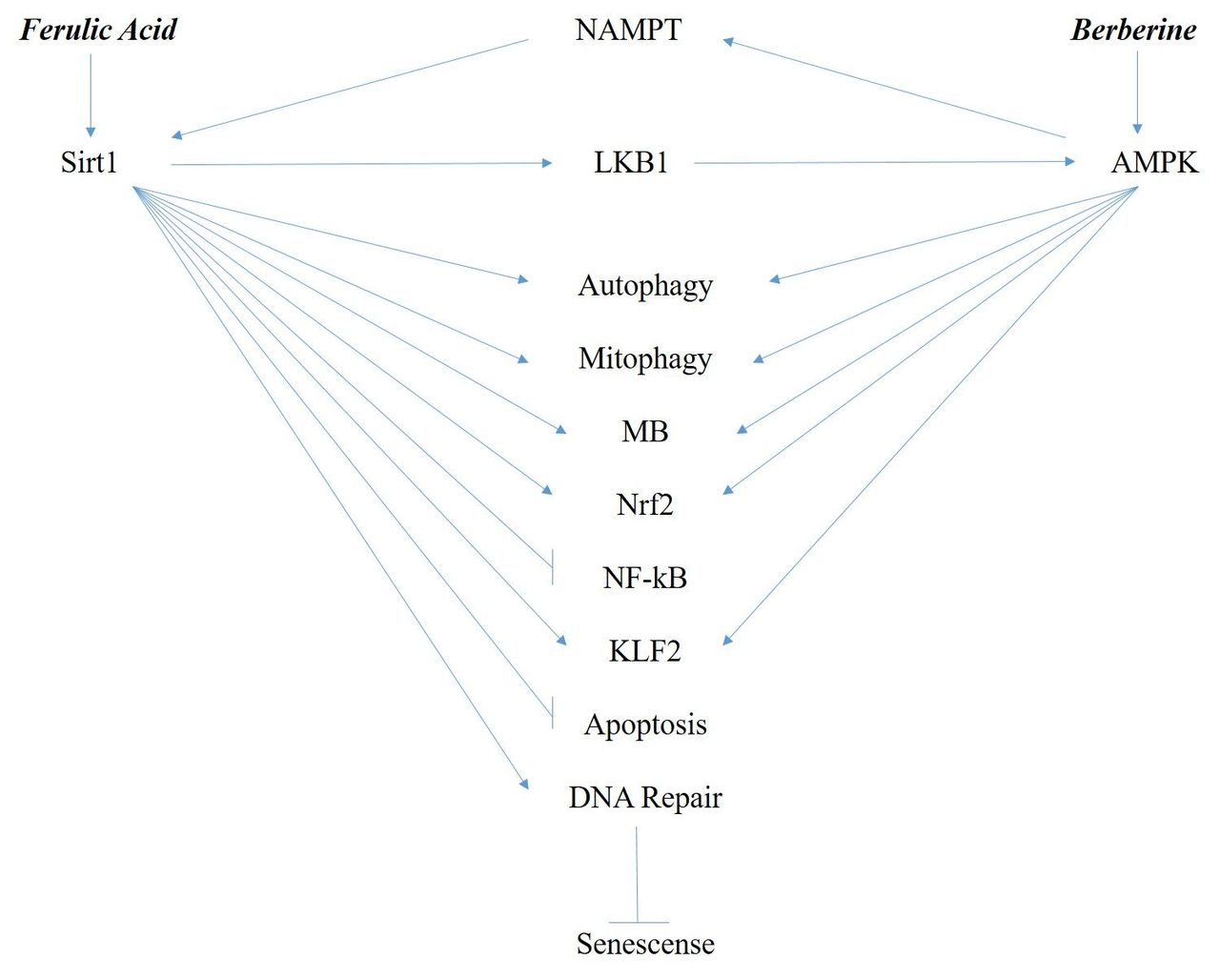
Figure 1Figure 1 summarizes the protective and often complementary effects of Sirt1 and AMPK on cellular processes, transcription factors, and other proteins.
{kind=link}
Some key complementary cell protective effects of Sirt1 and AMPK, as discussed earlier. AMPK, AMP-activated kinase; KLF2, kruppel-like factor 2; MB, mitochondrial biogenesis; NAMPT, nicotinamide phosphoribosyltransferase.
Ancillary nutraceuticals may amplify the beneficial impacts of ferulic acid/berberine
Melatonin can also cooperate with ferulic acid. Likely acting via its membrane receptors and the Bmal1 transcription factor, melatonin has been reported to upregulate the expression of both Nrf2 and Sirt1.91–96 Hence, a regimen of ferulic acid, metformin/berberine and melatonin could be expected to collaborate in promoting strong activation of Sirt1, AMPK and Nrf2—activities linked to health protection and longevity97–99—while downregulating the proinflammatory actions of NF-kappaB.
Activation of Nrf2 could be further boosted with nutraceuticals that disrupt Nrf2’s interaction with Keap1, such as lipoic acid and broccoli sprout extracts (which give rise to sulforaphane).100–102 Additionally, the spirulina chromophore phycocyanobilin may upregulate Nrf2 expression at the transcriptional level, possibly via agonism for the aryl hydrocarbon receptor.103–106
Nicotinamide riboside (NR), now available as a nutraceutical, can be transported into cells and phosphorylated to nicotinamide riboside-5-phosphate, the product of NAMPT. Hence, it, like AMPK activators, has potential to enhance the availability Sirt1’s substrate NAD+.107 108 Unfortunately, clinical supplementation studies involving NR intakes of up to 2 g daily have failed to show an impact of such supplementation on skeletal muscle NAD+ levels or indices of mitochondrial function; nor has NR supplementation so far shown an impact on insulin sensitivity in obese, insulin-resistant men.109 However, an increase of NAD+ in blood mononuclear cells has been reported in subjects receiving long-term NR supplementation at 500 mg two times a day.110 Hence, further clinical exploration of this agent appears warranted.
With respect to Sirt1, both its activity and its expression can be increased by endogenously generated hydrogen sulfide (H2S), which directly sulfhydrates Sirt1.111 This might rationalise several reports that supplemental taurine can boost Sirt1 activity; taurine has been shown to enhance expression of the H2S-generating enzymes cystathionine β-synthase and cystathionine γ-lyase in vascular tissues, an effect which might be generalisable to other tissues.112–116 Likewise, administration of N-acetylcysteine, a delivery form of the cysteine that serves as substrate for H2S biosynthesis, has been found to increase Sirt1 activity or expression in rodents.117 118
In curious parallel to ferulic acid, urolithin A is a bacterial metabolite of ellagitannins and ellagic acid (found in pomegranates, berries and nuts) thought to mediate the physiological effects of these poorly absorbed phytochemicals.119–121 In further parallel, urolithin A has been shown to promote mitophagy and MB in cell cultures and rodents; moreover, a similar effect has now been reported in the skeletal muscle of humans ingesting 500–1000 mg urolithin A daily.122–125 In light of these findings, a new report that urolithin A can enhance Sirt1 activity is not surprising.126 127 Urolithin A has very recently become available as a nutraceutical, and may have considerable promise for health promotion.122
PGC-1α collaborates with the PPARα transcription factor in promoting transcription of genes coding for various mitochondrial proteins that enable fatty acid oxidation and ketogenesis, such as the carnitine palmitoyltransferases, acyl-CoA oxidase and acetyl-CoA oxidase.128 129 The phytochemical antioxidant astaxanthin can function as a PPARα agonist in concentrations achievable via supplementation.130 131 This provides a likely explanation for the favourable effects of supplemental astaxanthin on aspects of metabolic syndrome, as reported in obese rodents and humans.132–135
NO, acting via cGMP and enhanced PGC-1α activity, has been found to support MB in skeletal muscle and other tissues.136 This may reflect cGMP/PKG1α-mediated activation of p38 MAP kinase via MEK3/6.137 138 p38 upregulates PGC-1α expression at the transcriptional level, and confers multiple phosphorylations on PGC-1α which increase its stability.43 Hence, agents which support or mimic NO synthase bioactivity may complement Sirt1’s impact on MB. In certain contexts, these may include citrulline, high-dose folate, dietary nitrate and high-dose biotin; the latter can act as a direct activator of soluble guanylate cyclase.139–141
Supplemental carnitine or acetylcarnitine has been shown to enhance MB in skeletal muscle, brain and liver of ageing rodents—an effect associated with increased expression of PGC-1α at the mRNA and protein level.142–145 This effect is not seen in younger animals, likely because supplemental carnitine has little effect on their tissue levels of carnitine. In contrast, carnitine levels in these tissues decline with age—likely owing to decreased expression of the carnitine transporter OCTN2—and carnitine supplementation restores more youthful tissue carnitine levels in old rodents.146–148 It has been suggested that the impact of supplemental carnitine on PGC-1α in ageing rodents reflect the ability of carnitine to moderately inhibit type 1 histone deacetylase activity in physiological tissue concentrations; the type 1 histone deacetylase HDAC3 activity has been reported to suppress PGC-1α expression at the transcriptional level.149–152 A clinical correlate of these findings is that, in elderly subjects complaining of ‘tiredness’, carnitine supplementation has been found in double-blind studies to boost perceived physical and mental energy.153–155
Because a ferulic acid/berberine regimen boosts induction of antioxidant enzymes via Nrf2 and FOXO transcription factors, while concurrently controlling mitochondrial oxidant production by stimulating mitophagy/MB, it can be viewed as a fruitful strategy for control of oxidative stress. Ancillary nutraceutical strategies in that regard include astaxanthin—which protects the structural integrity of the mitochondrial electron transport chain from oxidant damage156; phycocyanobilin—a spirulina chromophore which mimics free bilirubin in inhibiting certain forms of NADPH oxidase157 158; citrulline and high-dose folate—which promote recoupling of NO synthase in certain contexts139–141 159; and N-acetylcysteine, which can amplify glutathione synthesis.160 161
It should also be noted that exercise training can boost MB in the trained muscles. p38, along with calcium/calmodulin-dependent kinase II (CaMKII), upregulate PGC-1α activity and MB in exercised skeletal muscle.162 163 Oxidant production, primarily from xanthine oxidase, drives exercise-induced p38 activation, whereas activation of CaMKII is mediated by increased cytosolic free calcium.164
Conclusion
In summation, combined supplementation with ferulic acid and berberine, via mutually reinforcing activations of Sirt1 and AMPK, has the potential to promote a number of processes that help to keep cells ‘clean’ and properly functional—autophagy, mitophagy, MB, DNA repair, phase 2 induction—while aiding survival of non-renewable cells via suppression of apoptosis, senescence and NF-κB-driven inflammation. Exploration of this strategy in multiple rodent models of pathologies would be appropriate. Clinically, dose schedules of ferulic acid and of berberine that have shown physiological effects are in the range of 250–500 mg two times a day, and 500 mg, two/three times a day, respectively.
Data availability statement
No data are available.
Ethics statements
Patient consent for publication
Ethics approval
Not applicable.
References
Footnotes
Contributors All authors contributed to the final manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests JJD is Director of Scientific Affairs at Advanced Ingredients for Dietary Products. MM is coinventor and co-owner on a US patent for nutraceutical uses of phycocyanobilin oligopeptides derived from spirulina. SA and LMLL have no conflicts. JOK is an owner of a nutraceutical company.
Provenance and peer review Not commissioned; internally peer reviewed.