Article Text

Abstract
The α-glucosidase inhibitor acarbose, which slows carbohydrate digestion and blunts postprandial rises in plasma glucose, has long been used to treat patients with type 2 diabetes or glucose intolerance. Like metformin, acarbose tends to aid weight control, postpone onset of diabetes and decrease risk for cardiovascular events. Acarbose treatment can favourably affect blood pressure, serum lipids, platelet aggregation, progression of carotid intima-media thickness and postprandial endothelial dysfunction. In mice, lifetime acarbose feeding can increase median and maximal lifespan—an effect associated with increased plasma levels of fibroblast growth factor 21 (FGF21) and decreased levels of insulin-like growth factor-I (IGF-I). There is growing reason to suspect that an upregulation of fasting and postprandial production of glucagon-like peptide-1 (GLP-1)—stemming from increased delivery of carbohydrate to L cells in the distal intestinal tract—is largely responsible for the versatile health protection conferred by acarbose. Indeed, GLP-1 exerts protective effects on vascular endothelium, the liver, the heart, pancreatic β cells, and the brain which can rationalise many of the benefits reported with acarbose. And GLP-1 may act on the liver to modulate its production of FGF21 and IGF-I, thereby promoting longevity. The benefits of acarbose are likely mimicked by diets featuring slowly-digested ‘lente’ carbohydrate, and by certain nutraceuticals which can slow carbohydrate absorption. Prebiotics that promote colonic generation of short-chain fatty acids represent an alternative strategy for boosting intestinal GLP-1 production. The health benefits of all these measures presumably would be potentiated by concurrent use of dipeptidyl peptidase 4 inhibitors, which slow the proteolysis of GLP-1 in the blood.
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Acarbose therapy promotes vascular health
Acarbose is an α-glucosidase inhibitor prescribed for prevention and treatment of diabetes; it and metformin are the most commonly used diabetes drugs worldwide. Less than 2% of acarbose is absorbed after administration, rationalising its non-toxicity; its clinical utility evidently reflects its ability to slow absorption of dietary carbohydrate by acting within the intestinal tract to inhibit brush-border α-glucosidase, which plays a key role in the digestion of starch and sugars.1 ,2 Inhibition of α-glucosidase in saliva, and non-competitive inhibition of pancreatic α-amylase, also contribute to it slowing of carbohydrate absorption.3 Clinical doses of acarbose ingested with meals notably blunt the postprandial increase in serum glucose, a well-documented predictor of macrovascular and microvascular complications in diabetics, and of macrovascular risk in non-diabetics.4 ,5 A Cochrane meta-analysis found that, on average, acarbose therapy lowers glycated haemoglobin (HbA1c) levels by 0.8%, and reduces the postprandial increase in plasma glucose by 2.3 mM.6 Glycaemic control benefits of acarbose therapy tend to be greater in Asian clinical trials than in Western ones, likely because Asians tend to consume diets higher in carbohydrates.7 Gastrointestinal side effects of acarbose—reflecting carbohydrate malabsorption—are fairly common when initiating therapy, but tend to lessen over time as the distal intestine upregulates its capacity to absorb glucose. These side effects can be minimised by starting with a low dose—25 or 50 mg with two or three meals daily—and gradually raising it; in other words, ‘start low, go slow’.8 The maximal dose of acarbose is 100 mg three time a day—higher doses do not achieve a greater impact on carbohydrate digestion.
Like metformin, but unlike many other diabetes drugs, acarbose tends to promote weight loss, and cannot trigger hypoglycaemic episodes.9–11 Acarbose also resembles metformin in that there is reasonably compelling evidence that acarbose therapy in patients with diabetes or glucose intolerance has a favourable impact on cardiovascular health outcomes, and can slow the onset of diabetes in glucose-intolerant participants.2 ,12 ,13 Analyses of long-term outcomes in the Stop Non-insulin-dependent Diabetes Mellitus (STOP-NIDDM) trial, as well as a meta-analysis of longer-term controlled trials of acarbose in type 2 diabetics, conclude that risk for cardiovascular events, most notably myocardial infarction, is lower in acarbose-treated patients (HR=0.51; 95% CI 0.28 to 0.95 in STOP-NIDDM and HR=0.66; CI 0.48 to 0.88 in the diabetes treatment meta-analysis).14 ,15 A large multicenter controlled trial in China ongoing since 2009, the Acarbose Cardiovascular Evaluation study, should provide a more definitive determination of acarbose's capacity to promote vascular health.16
Acarbose therapy can favourably influence cardiovascular risk factors. In diabetics, acarbose therapy tends to lower mean blood pressure; in the STOP-NIDDM trial, incidence of newly diagnosed hypertension was 34% lower in the acarbose-treated group.14 ,17 Yet acarbose therapy also helps to prevent postprandial hypotension in elderly participants prone to this disorder.18 ,19 Serum triglyceride levels tend to drop, and high-density lipoprotein cholesterol to rise, during acarbose therapy.13 ,14 In early type 2 diabetes, acarbose therapy decreases urinary markers of platelet aggregation.20 Slowed progression of carotid intima-media thickness has also been reported in acarbose-treated patients, and acarbose therapy improves postprandial endothelium-dependent vasodilation.21–24 In rabbits subjected to 30 min of coronary occlusion followed by reperfusion, feeding acarbose for a week prior to the procedure was associated with a greater than 50% reduction in infarct size; this benefit was wholly abolished if an inhibitor of ATP-sensitive K channels (5-hydroxydecanoate) was administered slightly before onset of ischaemia.25
Acarbose extends longevity and modulates FGF21 and insulin-like growth factor-I (IGF-I) in mice
There is now reason to suspect that, beyond its clear utility in those who are glucose intolerant, acarbose may have broader potential for health promotion. In a lifetime feeding study in mice, addition of 0.1% acarbose to a standard diet (65% of calories from grain carbohydrates, 22% protein), beginning at 4 months of age, was associated with significant increases in the median and maximal lifespan, in males and females.26 The increase in median lifespan was greater in males than females—22% vs 5%; maximal lifespan increased 11% and 9% in males and females, respectively. Curiously, acarbose feeding did not influence the HbA1c level in these mice, likely because their fasting glucoses were slightly higher, balancing out the likely impact of acarbose on postprandial glucose. Since these mice were not diabetic or diabetes prone, it is not immediately apparent why slowing dietary glucose absorption would have such a notable impact on longevity in mice.
However, the researchers uncovered two key clues—serum FGF21 was significantly higher, and serum insulin-like growth factor-I (IGF-I) significantly lower in the acarbose-treated mice.26 Systemic IGF-I activity is a clear driver of the aging process and determinant of longevity in mice;27 moreover, FGF21 acts on the liver to blunt hepatic sensitivity to growth hormone, thereby decreasing the liver's production of IGF-I.28–30 Transgenic mice with a constitutive increase in FGF21 production enjoy a substantial increase in mean and maximal lifespan, an effect likely mediated in part by downregulation of IGF-I activity.31 ,32
An effect of acarbose on hepatic FGF21 production has not previously been reported. However, by diverting additional glucose to the distal intestine, chronic therapy with acarbose or other α-glucosidase inhibitors boosts the production of glucagon-like peptide-1 (GLP-1) by intestinal L cells.33–38 There are several reports that long-acting agonists for the GLP-1 receptor increase hepatic FGF21 production.39 ,40 PPARα, after deacetylation by Sirt1, interacts with the FGF21 promoter to stimulate transcription of this hormone.41–45 Notably, GLP-1 agonist drugs are reported to increase expression of PPARα and Sirt1 in hepatocytes.46–48 Hence, these findings suggest that the upregulation of GLP-1 consequent to chronic acarbose treatment may act on the liver to promote transcriptional activation of FGF21, which in turn suppresses IGF-I production.
Acarbose benefits reflect suppression of postprandial glycaemia and upregulation of GLP-1
In light of current evidence, two phenomena appear to interact in mediating acarbose's favourable health impacts—a suppression of postprandial surges in plasma glucose, and an increased production of GLP-1. A rapid elevation of plasma glucose levels imposes an oxidative stress on vascular endothelium that compromises the protective function of endothelial nitric oxide synthase (eNOS) and promotes inflammation; this effect is associated with an acute reduction in endothelium-dependent vasodilation.49 This oxidative stress can originate from NADPH oxidase, uncoupled eNOS, and mitochondria.50–52 Curiously, episodic large rises in glucose appear to have a more notable impact in this regard than sustained elevations of glucose; hence, blunting the postprandial glucose rise may be particularly worthwhile as a strategy for protecting the vasculature from oxidative stress.49 ,53 ,54 Moreover, since acarbose therapy does not risk provoking hypoglycaemic reactions, it might be viewed as a strategy for stabilising glucose levels in diabetics, avoiding the wild swings in glycaemia that may be particularly noxious for vascular health. As noted, postprandial glucose predicts risk for microvascular and macrovascular complications in type 2 diabetics more effectively than fasting glucose; it also predicts vascular risk in glucose-intolerant non-diabetics.4 ,5 While some of this risk likely is attributable to metabolic factors that induce this glucose intolerance, there is also good reason to suspect that the pro-oxidative impact of postprandial hyperglycaemia per se is a key mediator of this risk. Moreover, episodic hyperglycaemia, in conjunction with elevated free fatty acids, also exerts an oxidant effect on pancreatic β cells; in pre-diabetics, this ‘glucolipotoxicity’ can lead to the failure of glucose-stimulated insulin secretion and β-cell apoptosis that ushers in clinical diabetes.55 Hence, moderation of postprandial hyperglycaemia may be largely responsible for acarbose's ability to slow diabetes onset.
However, the ability of acarbose treatment to up-regulate GLP-1 production may also be a key mediator of its health benefits. A recent 24-week study in type 2 diabetics found that fasting and postprandial plasma concentrations of GLP-1 were about 10% and 20% higher, respectively, during acarbose therapy.38 GLP-1 acts directly on vascular endothelium to boost eNOS activity;56–58 in the 24-week study, changes in GLP-1 levels correlated directly with increases in serum nitric oxide.38 Activation of AMPK also contributes to GLP-1's favourable influence on endothelial function.59 GLP-1 acts directly on the liver to promote fatty acid oxidation and inhibit fatty acid synthesis;47 ,60 GLP-1 receptor agonists have a favourable impact on models of non-alcoholic fatty liver disease in rodents, and also have a favourable effect on this syndrome in humans.46 ,61–64 The clinical impact of acarbose on non-alcoholic fatty liver disease still requires evaluation.65 As we have seen, the effects of acarbose treatment on hepatic production of FGF21 and IGF-I in healthy mice might well be attributable to increased GLP-1 production.
The tendency of acarbose to promote weight loss is shared by long-lasting GLP-1 receptor mimics currently used in diabetes therapy; indeed, these agents can aid weight loss in non-diabetics as well.66 ,67 Hence, it is reasonable to suspect that GLP-1 is a mediator of the favourable impact of acarbose therapy on weight control.
Studies with long-lived GLP-1 agonists in rodents demonstrate that pretreatment with these agents ameliorate the impact of ischaemia reperfusion on the heart.68–72 This protective effect is blocked by concurrent administration of inhibitors of mitochondrial ATP-sensitive K channels (mKATP).72 Ischaemic preconditioning likewise protects the heart from ischaemia-reperfusion by up-regulating activity of mKATP channels.73 The opening of these channels during ischaemia lessens the potential gradient driving calcium influx into mitochondria, preventing an excessive rise in intramitochondrial calcium that severely compromises mitochondrial function.39 ,47 The fact that acarbose pretreatment of normal healthy rabbits markedly decreases infarct size following myocardial ischaemia reperfusion—a benefit blocked by acute administration of an mKATP inhibitor—suggests that, by raising GLP-1 production, acarbose exerts a protective effect on the heart mechanistically analogous to ischaemic preconditioning.25
In addition to acting directly on pancreatic β cells to potentiate glucose-stimulated insulin release, GLP-1 also acts on these cells in various ways to counteract glucolipotoxicity.74–78 Hence, increased GLP-1 production may collaborate with a decrease in postprandial glucose to mediate acarbose's diabetes preventive action.
Brain neurons express GLP-1 receptors, and rodent studies with long-lived GLP-1 receptor agonists indicated that these agents can protect neurons from ischaemic damage, support effective learning, and diminish memory impairment in mouse models of Alzheimer's disease.79–86 Since these agents are effective in these regards when administered intraperitoneally, it is conceivable that chronic upregulation of intestinal GLP-1 production could modestly benefit brain health.
In summary, GLP-1 acts directly on the vasculature, liver, myocardium, β cells, and brain to safeguard the structural and functional integrity of these organs—while aiding weight control and modulating FGF21 and IGF-I in a way likely to slow aging and promote longevity; these interactions are summarised in figure 1. Acarbose therapy, or other measures which safely up-regulate intestinal GLP-1 production, may provide protection in these respects. And there is no reason to believe that such benefits will only be achievable in the context of glucose intolerance; upregulation of GLP-1 production might be viewed as an ‘anti-aging’ strategy of benefit to the general population. Of course, acarbose is likely to provide greater net protection in glucose-intolerant participants, owing to its favourable influence on their excessive postprandial glucose excursions.
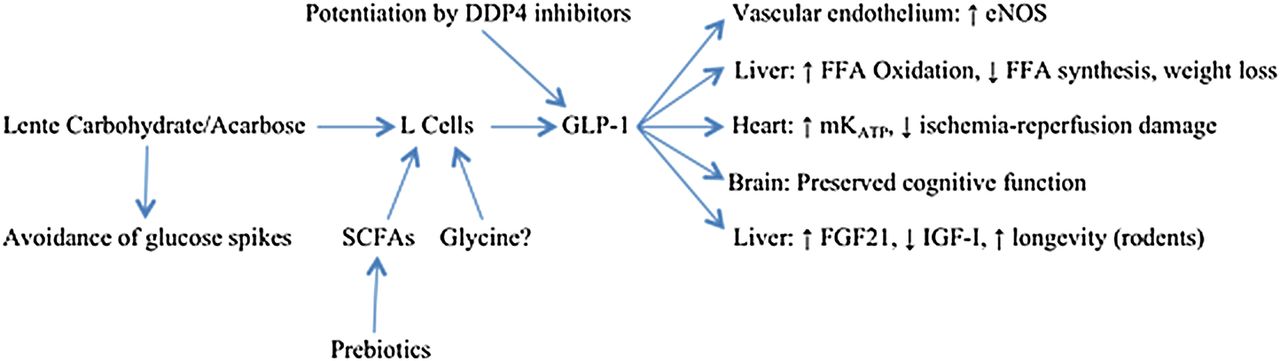
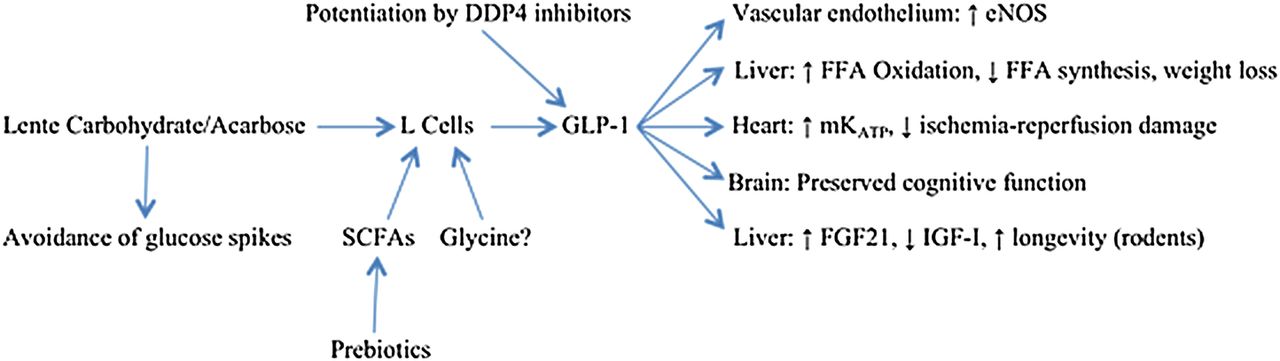{kind=link}
Potential health benefits of lente carbohydrate/acarbose. GLP, glucagon-like peptide-1; FGF21, fibroblast growth factor 21; IGF-I, insulin-like growth factor-I; eNOS, endothelial nitric oxide synthase; FFA, free fatty acids; SCFA, short-chain fatty acids.
Alternative or complementary strategies for achieving analogous health benefits
Since acarbose, for practical purposes, is not absorbed, and its physiological effects hence reflect a slowing of dietary carbohydrate digestion, it is reasonable to suspect that choosing a diet in which slow-digesting ‘lente’ carbohydrate predominates should achieve benefits comparable to those of acarbose therapy.87 ,88 Pasta, boiled beans, structurally intact grain kernels and sprouted grains—as contrasted to most flour or sugar products—are more difficult to digest and have a notably lower glycaemic index.89 Arguably, complementing such dietary choices with a low dose of acarbose (perhaps 25 mg per meal?) would be an appropriate strategy for minimising postprandial glycaemic excursion while boosting GLP-1 production.
Nutraceutical measures for delaying digestion of dietary carbohydrate, including administration of adequate doses of the highly viscous soluble fibre glucomannan, or of amylase-inhibitory phytochemicals extracted from beans, might be expected to mimic the health benefits achievable by acarbose or other pharmaceutical α-glucosidase inhibitors (eg, miglitol, voglibose).90–98 Glucomannan has the potential drawback of impeding the absorption of certain coadministered drugs or fat-soluble vitamins; hence, its use may be incompatible with some drugs that must be administered with meals.99 ,100 Prebiotics can promote colonic production of GLP-1, as the short-chain fatty acids produced by properly nourished gut bacteria act on L cells to stimulate release of GLP-1.101–106 Hence, prebiotics such as inulin or resistant starch may mimic the GLP-1-mediated benefits of acarbose, though they could not be expected to notably influence postprandial glucose levels. Consideration should also be given to the possibility that dietary glycine might act on L cells to stimulate GLP-1 release; high physiological concentrations of glycine, via activation of glycine-gated chloride channels, have this effect on an L-cell-derived cell line.107 Such an effect might explain the ability of glycine supplementation to prevent metabolic syndrome and fatty liver in sucrose-fed rats.108–110
The up-regulatory impact of any of these measures on GLP-1 levels should be considerably potentiated by concurrent administration of dipeptidyl peptidase 4 (DPP4) inhibitors—for example, sitagliptin, vildagliptin—which slow the proteolytic degradation of GLP-1 in plasma.111
In overview, it appears that achieving slow digestion of dietary carbohydrate—either by choosing ‘lente’ carbohydrate foods, or by pharmacological inhibition of carbohydrate digestion, as with acarbose—can exert a range of favourable effects on health and longevity by up-regulating GLP-1 production and concurrently blunting the pro-oxidative effect of postprandial glucose elevations. Prebiotics can also be employed to boost GLP-1 production, and concurrent treatment with DPP4 inhibitors should potentiate the impact of these strategies on GLP-1 levels. These strategies may have potential for optimising healthspan not just in diabetics and pre-diabetics, but in the general population.
References
Footnotes
Contributors MFM and JJDN are solely responsible for the conception and writing of this review.
Competing interests MFM is owner and science director of the small nutraceutical company NutriGuard Research, which sells glucomannan.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement No additional data are available.